

What Is The Main Function Of The Golgi In An Animal Cell?
The Golgi apparatus, or Golgi complex, functions as a factory in which proteins received from the ER are farther processed and sorted for transport to their eventual destinations: lysosomes, the plasma membrane, or secretion. In addition, every bit noted before, glycolipids and sphingomyelin are synthesized within the Golgi. In constitute cells, the Golgi apparatus further serves as the site at which the complex polysaccharides of the cell wall are synthesized. The Golgi apparatus is thus involved in processing the broad range of cellular constituents that travel along the secretory pathway.
Organisation of the Golgi
Morphologically the Golgi is equanimous of flattened membrane-enclosed sacs (cisternae) and associated vesicles (Figure 9.22). A striking feature of the Golgi appliance is its distinct polarity in both structure and office. Proteins from the ER enter at its cis confront (entry face), which is convex and usually oriented toward the nucleus. They are then transported through the Golgi and exit from its concave trans face up (exit face). As they pass through the Golgi, proteins are modified and sorted for transport to their eventual destinations within the cell.

Figure 9.22
Electron micrograph of a Golgi apparatus. The Golgi apparatus consists of a stack of flattened cisternae and associated vesicles. Proteins and lipids from the ER enter the Golgi apparatus at its cis face and exit at its trans confront. (Courtesy of Dr. L. (more...)
Singled-out processing and sorting events appear to take place in an ordered sequence inside different regions of the Golgi complex, so the Golgi is unremarkably considered to consist of multiple discrete compartments. Although the number of such compartments has non been established, the Golgi is nigh normally viewed as consisting of iv functionally distinct regions: the cis Golgi network, the Golgi stack (which is divided into the medial and trans subcompartments), and the trans Golgi network (Figure ix.23). Proteins from the ER are transported to the ER-Golgi intermediate compartment and then enter the Golgi apparatus at the cis Golgi network. They then progress to the medial and trans compartments of the Golgi stack, inside which most metabolic activities of the Golgi appliance accept place. The modified proteins, lipids, and polysaccharides then motility to the trans Golgi network, which acts as a sorting and distribution center, directing molecular traffic to lysosomes, the plasma membrane, or the cell exterior.

Figure 9.23
Regions of the Golgi appliance. Vesicles from the ER fuse to form the ER-Golgi intermediate compartment, and proteins from the ER are and then transported to the cis Golgi network. Resident ER proteins are returned from the ER-Golgi intermediate compartment (more...)
Although the Golgi apparatus was first described over 100 years agone, the mechanism past which proteins movement through the Golgi apparatus has nonetheless not been established and is an surface area of controversy amongst cell biologists. 1 possibility is that transport vesicles deport proteins between the cisternae of the Golgi compartments. However, in that location is considerable experimental support for an alternative model proposing that proteins are simply carried through compartments of the Golgi within the Golgi cisternae, which gradually mature and progressively move through the Golgi in the cis to trans management.
Protein Glycosylation inside the Golgi
Poly peptide processing within the Golgi involves the modification and synthesis of the carbohydrate portions of glycoproteins. 1 of the major aspects of this processing is the modification of the Northward-linked oligosaccharides that were added to proteins in the ER. As discussed earlier in this affiliate, proteins are modified within the ER by the addition of an oligosaccharide consisting of xiv sugar residues (see Figure 9.15). Iii glucose residues and i mannose are so removed while the polypeptides are yet in the ER. Following send to the Golgi apparatus, the N-linked oligosaccharides of these glycoproteins are subject to all-encompassing further modifications.
N-linked oligosaccharides are candy within the Golgi apparatus in an ordered sequence of reactions (Figure nine.24). The first modification of proteins destined for secretion or for the plasma membrane is the removal of iii additional mannose residues. This is followed by the sequential addition of an Due north-acetylglucosamine, the removal of two more mannoses, and the add-on of a fucose and two more than N-acetylglucosamines. Finally, iii galactose and iii sialic acid residues are added. Equally noted in Chapter 7, different glycoproteins are modified to different extents during their passage through the Golgi, depending on both the structure of the protein and on the amount of processing enzymes that are present within the Golgi complexes of dissimilar types of cells. Consequently, proteins can emerge from the Golgi with a variety of different N-linked oligosaccharides.

Figure 9.24
Processing of Due north-linked oligosaccharides in the Golgi. The Northward-linked oligosaccharides of glycoproteins transported from the ER are farther modified by an ordered sequence of reactions in the Golgi.
The processing of the N-linked oligosaccharide of lysosomal proteins differs from that of secreted and plasma membrane proteins. Rather than the initial removal of three mannose residues, proteins destined for incorporation into lysosomes are modified by mannose phosphorylation. In the showtime step of this reaction, N-acetylglucosamine phosphates are added to specific mannose residues, probably while the protein is still in the cis Golgi network (Figure 9.25). This is followed past removal of the Northward-acetylglucosamine group, leaving mannose-six-phosphate residues on the N-linked oligosaccharide. Because of this modification, these residues are not removed during further processing. Instead, these phosphorylated mannose residues are specifically recognized by a mannose-6-phosphate receptor in the trans Golgi network, which directs the transport of these proteins to lysosomes.

Effigy nine.25
Targeting of lysosomal proteins by phosphorylation of mannose residues. Proteins destined for incorporation into lysosomes are specifically recognized and modified by the add-on of phosphate groups to the half-dozen position of mannose residues. In the first (more...)
The phosphorylation of mannose residues is thus a critical pace in sorting lysosomal proteins to their correct intracellular destination. The specificity of this process resides in the enzyme that catalyzes the first step in the reaction sequence—the selective add-on of N-acetylglucosamine phosphates to lysosomal proteins. This enzyme recognizes a structural determinant that is nowadays on lysosomal proteins merely not on proteins destined for the plasma membrane or secretion. This recognition determinant is non a simple sequence of amino acids; rather, it is formed in the folded protein by the juxtaposition of amino acid sequences from different regions of the polypeptide chain. In contrast to the signal sequences that straight protein translocation to the ER, the recognition determinant that leads to mannose phosphorylation, and thus ultimately targets proteins to lysosomes, depends on the three-dimensional conformation of the folded protein. Such determinants are called signal patches, in contrast to the linear targeting signals discussed before in this chapter.
Proteins can likewise be modified by the add-on of carbohydrates to the side bondage of acceptor serine and threonine residues within specific sequences of amino acids (O-linked glycosylation) (run into Figure 7.28). These modifications take place in the Golgi apparatus past the sequential addition of single sugar residues. The serine or threonine is usually linked directly to Due north-acetylgalactosamine, to which other sugars can then exist added. In some cases, these sugars are farther modified by the improver of sulfate groups.
Lipid and Polysaccharide Metabolism in the Golgi
In add-on to its activities in processing and sorting glycoproteins, the Golgi apparatus functions in lipid metabolism—in particular, in the synthesis of glycolipids and sphingomyelin. Every bit discussed earlier, the glycerol phospholipids, cholesterol, and ceramide are synthesized in the ER. Sphingomyelin and glycolipids are so synthesized from ceramide in the Golgi apparatus (Figure ix.26). Sphingomyelin (the only nonglycerol phospholipid in cell membranes) is synthesized past the transfer of a phosphorylcholine grouping from phosphatidylcholine to ceramide. Alternatively, the add-on of carbohydrates to ceramide can yield a diverseness of different glycolipids.
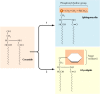
Figure 9.26
Synthesis of sphingomyelin and glycolipids. Ceramide, which is synthesized in the ER, is converted either to sphingomyelin (a phospholipid) or to glycolipids in the Golgi appliance. In the first reaction, a phosphorylcholine group is transferred from (more...)
Sphingomyelin is synthesized on the lumenal surface of the Golgi, but glucose is added to ceramide on the cytosolic side. Glucosylceramide then obviously flips, however, and additional carbohydrates are added on the lumenal side of the membrane. Neither sphingomyelin nor the glycolipids are so able to translocate across the Golgi membrane, so they are plant only in the lumenal half of the Golgi bilayer. Following vesicular transport, they are correspondingly localized to the exterior one-half of the plasma membrane, with their polar caput groups exposed on the prison cell surface. Every bit will exist discussed in Affiliate 12, the oligosaccharide portions of glycolipids are of import surface markers in cell-cell recognition.
In establish cells, the Golgi apparatus has the additional task of serving as the site where complex polysaccharides of the cell wall are synthesized. As discussed further in Affiliate 12, the plant cell wall is composed of three major types of polysaccharides. Cellulose, the predominant constituent, is a simple linear polymer of glucose residues. It is synthesized at the cell surface by enzymes in the plasma membrane. The other cell wall polysaccharides (hemicelluloses and pectins), even so, are complex, branched chain molecules that are synthesized in the Golgi apparatus and and so transported in vesicles to the cell surface. The synthesis of these cell wall polysaccharides is a major cellular function, and as much as eighty% of the metabolic activity of the Golgi appliance in institute cells may be devoted to polysaccharide synthesis.
Protein Sorting and Export from the Golgi Apparatus
Proteins, besides equally lipids and polysaccharides, are transported from the Golgi apparatus to their final destinations through the secretory pathway. This involves the sorting of proteins into unlike kinds of transport vesicles, which bud from the trans Golgi network and deliver their contents to the appropriate cellular locations (Effigy 9.27). Some proteins are carried from the Golgi to the plasma membrane by a constitutive secretory pathway, which accounts for the incorporation of new proteins and lipids into the plasma membrane, every bit well as for the continuous secretion of proteins from the cell. Other proteins are transported to the cell surface by a distinct pathway of regulated secretion or are specifically targeted to other intracellular destinations, such as lysosomes in fauna cells or vacuoles in yeast.

Figure ix.27
Transport from the Golgi appliance. Proteins are sorted in the trans Golgi network and transported in vesicles to their terminal destinations. In the absence of specific targeting signals, proteins are carried to the plasma membrane by constitutive secretion. (more...)
Proteins that part within the Golgi apparatus must exist retained within that organelle, rather than being transported along the secretory pathway. In contrast to the ER, all of the proteins retained inside the Golgi circuitous are associated with the Golgi membrane rather than beingness soluble proteins inside the lumen. The signals responsible for retentivity of some proteins within the Golgi have been localized to their transmembrane domains, which retain proteins within the Golgi appliance past preventing them from being packaged in the transport vesicles that get out the trans Golgi network. In addition, like the KKXX sequences of resident ER membrane proteins, signals in the cytoplasmic tails of some Golgi proteins mediate the retrieval of these proteins from subsequent compartments forth the secretory pathway.
The constitutive secretory pathway, which operates in all cells, leads to continual unregulated protein secretion. However, some cells also possess a distinct regulated secretory pathway in which specific proteins are secreted in response to environmental signals. Examples of regulated secretion include the release of hormones from endocrine cells, the release of neurotransmitters from neurons, and the release of digestive enzymes from the pancreatic acinar cells discussed at the beginning of this chapter (encounter Figure 9.2). Proteins are sorted into the regulated secretory pathway in the trans Golgi network, where they are packaged into specialized secretory vesicles. These secretory vesicles, which are larger than other transport vesicles, shop their contents until specific signals direct their fusion with the plasma membrane. For case, the digestive enzymes produced by pancreatic acinar cells are stored in secretory vesicles until the presence of food in the tum and minor intestine triggers their secretion. The sorting of proteins into the regulated secretory pathway appears to involve the recognition of signal patches shared by multiple proteins that enter this pathway. These proteins selectively aggregate in the trans Golgi network and are and so released by budding as secretory vesicles.
A farther complexity in the transport of proteins to the plasma membrane arises in many epithelial cells, which are polarized when they are organized into tissues. The plasma membrane of such cells is divided into two separate regions, the apical domain and the basolateral domain, that contain specific proteins related to their particular functions. For case, the apical membrane of intestinal epithelial cells faces the lumen of the intestine and is specialized for the efficient absorption of nutrients; the remainder of the jail cell is covered by the basolateral membrane (Figure ix.28). Distinct domains of the plasma membrane are nowadays not merely in epithelial cells, simply also in other cell types. Thus, the constitutive secretory pathway must selectively transport proteins from the trans Golgi network to these singled-out domains of the plasma membrane. This is accomplished by the selective packaging of proteins into at least 2 types of constitutive secretory vesicles that leave the trans Golgi network targeted specifically for either the apical or basolateral plasma membrane domains of the cell.
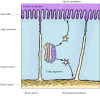
Figure nine.28
Transport to the plasma membrane of polarized cells. The plasma membranes of polarized epithelial cells are divided into upmost and basolateral domains. In this instance (intestinal epithelium), the apical surface of the cell faces the lumen of the intestine, (more...)
The all-time-characterized pathway of protein sorting in the Golgi is the selective transport of proteins to lysosomes. As already discussed, lumenal lysosomal proteins are marked past mannose-6-phosphates that are formed by modification of their N-linked oligosaccharides shortly later on entry into the Golgi apparatus. A specific receptor in the membrane of the trans Golgi network then recognizes these mannose-half-dozen-phosphate residues. The resulting complexes of receptor plus lysosomal enzyme are packaged into send vesicles destined for lysosomes. Lysosomal membrane proteins are targeted by sequences in their cytoplasmic tails, rather than by mannose-6-phosphates.
In yeasts and plant cells, which lack lysosomes, proteins are transported from the Golgi apparatus to an additional destination: the vacuole (Figure nine.29). Vacuoles assume the functions of lysosomes in these cells as well as performing a multifariousness of other tasks, such as the storage of nutrients and the maintenance of turgor pressure and osmotic rest. In contrast to lysosomal targeting, proteins are directed to vacuoles past short peptide sequences instead of carbohydrate markers.

Figure 9.29
A found jail cell vacuole. The big central vacuole functions as a lysosome in addition to storing nutrients and maintaining osmotic residue. (E. H. Newcombe/Biological Photo Service.)
Source: https://www.ncbi.nlm.nih.gov/books/NBK9838/
Posted by: herronoverniseents1972.blogspot.com
0 Response to "What Is The Main Function Of The Golgi In An Animal Cell?"
Post a Comment